阿部 文快
独立行政法人海洋研究開発機構 極限環境生物圏研究センター 代謝・適応機能研究グループ グループリーダー
横浜市立大学大学院国際総合科 学研究科 環境生命系 客員教授 博士(理学)
Readout No.34 May 2008
ライフサイエンス,特に生きた細胞を対象とする研究領域では,蛍光化合物を標識とした分子イメージングが現在盛んだ。これまで多くの興味は標的分子の量的変動や局在に向けられてきたが,最近,蛍光寿命や偏光解消といった蛍光分子が持つ魅力的な性質を利用するケースが増えてきている。本稿では,その基本的な原理と応用,及び蛍光寿命測定装置FluoroCubeを用いて行った著者らの研究例について,いくつか紹介したい。
現代ライフサイエンスは,“蛍光イメージング革命”と称されるほど飛躍的に向上した蛍光化合物の利用と,ハード面の充実に裏打ちされ,かつて想像すらし得なかった生き物の“なまの姿”を次々とあぶり出している。細胞内pHやCa2+濃度の変化に応答して蛍光強度比が変わるインジケータ,間接蛍光抗体法に用いる色とりどりの蛍光色素(Invitrogen社のAlexaシリーズなど),あるいは目的タンパク質との融合産物として細胞内で発現させ,ライブセルイメージングを可能にする緑色蛍光タンパク質(Green Fluorescent Protein:GFP)などがその代表例である。
ライフサイエンスでは,蛍光物質を用いた技法のほとんどが,細胞内で進行している眼に見えない現象を蛍光を標識に観察・計測するために行われている。目的タンパク質は細胞のどのあたりに局在しているのか?存在量は多いのか少ないのか?細胞に刺激を与えると,内部のpHやCa2+濃度はどう変化するのか?いずれも蛍光の“強度”が観察・計測の対象となる。一方,蛍光分子は“蛍光寿命”と“蛍光偏光”といった魅力的な特徴をあわせ持つ。筆者のいる細胞生物学・微生物学の分野でも,これらを積極的に活用しようとする動きが見られてきている。
蛍光寿命や蛍光偏光を知ることで細胞の何が理解されるのか?筆者は蛍光分光学の専門家ではないので,本稿ではアプリケーションに力を注ぐ現場の生物学者の立場から解説したい。実例として,HORIBA Jobin Yvon社のFluoroCubeを用いて得られた筆者らの最近の知見にも触れたい。なお,優れた書籍が出版されているので参考文献[1-3]として掲げている。
まず,ごく簡単に蛍光寿命の意味について説明する。
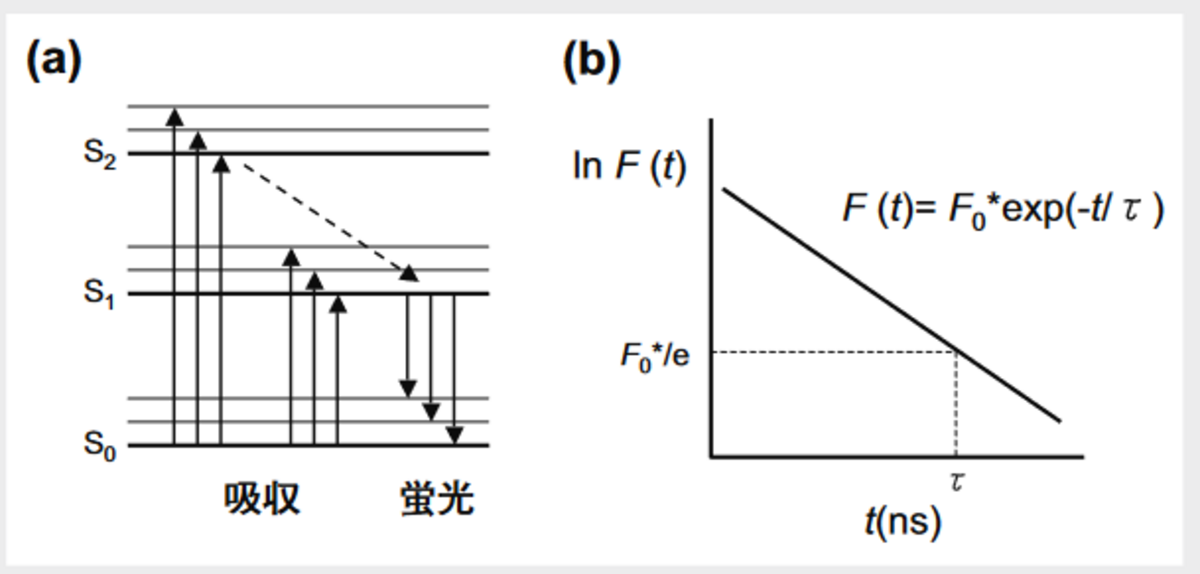
図1(a)に示すように,蛍光分子が基底状態にある時,電子はS0と呼ばれるエネルギー状態にある。蛍光分子が光エネルギーを吸収すると,わずかフェムト秒のオーダで励起状態に遷移し,過剰なエネルギーを散逸して第1励起状態S1の最低次の振動レベルまで落ちる。この状態は励起過程で最も安定であり,滞在時間は数十から数ナノ秒が普通である。ここから基底状態に戻る過程で蛍光が発する。蛍光強度の時間変化F(t)は式(1)で与えられる(図1(b))。
![]() (1)
(1)
ここで,F0はt=0における蛍光強度でτが蛍光寿命である。最初に励起状態にあった蛍光分子数に比べて,τだけ時間がたってもまだ励起状態にある蛍光分子数が1/e(約37%)になっていることになる(図1(b))。蛍光分子周辺の誘電率が高い時(親水的),あるいは近傍にエネルギー受容体があるとτは短くなる。従って,τを調べることで蛍光分子の周りがどんな環境にあるのかが理解される。
図1 蛍光分子の電子状態と蛍光の減衰曲線 (a)蛍光分子の電子状態:基底状態(S₀)にある分子は励起エネルギーを吸収し,励起状態S₁やS₂に遷移する。最低次の励起状態S₁から基底状態に遷移するとき蛍光が発する。 (b)蛍光の減衰曲線:τは蛍光寿命を示す。
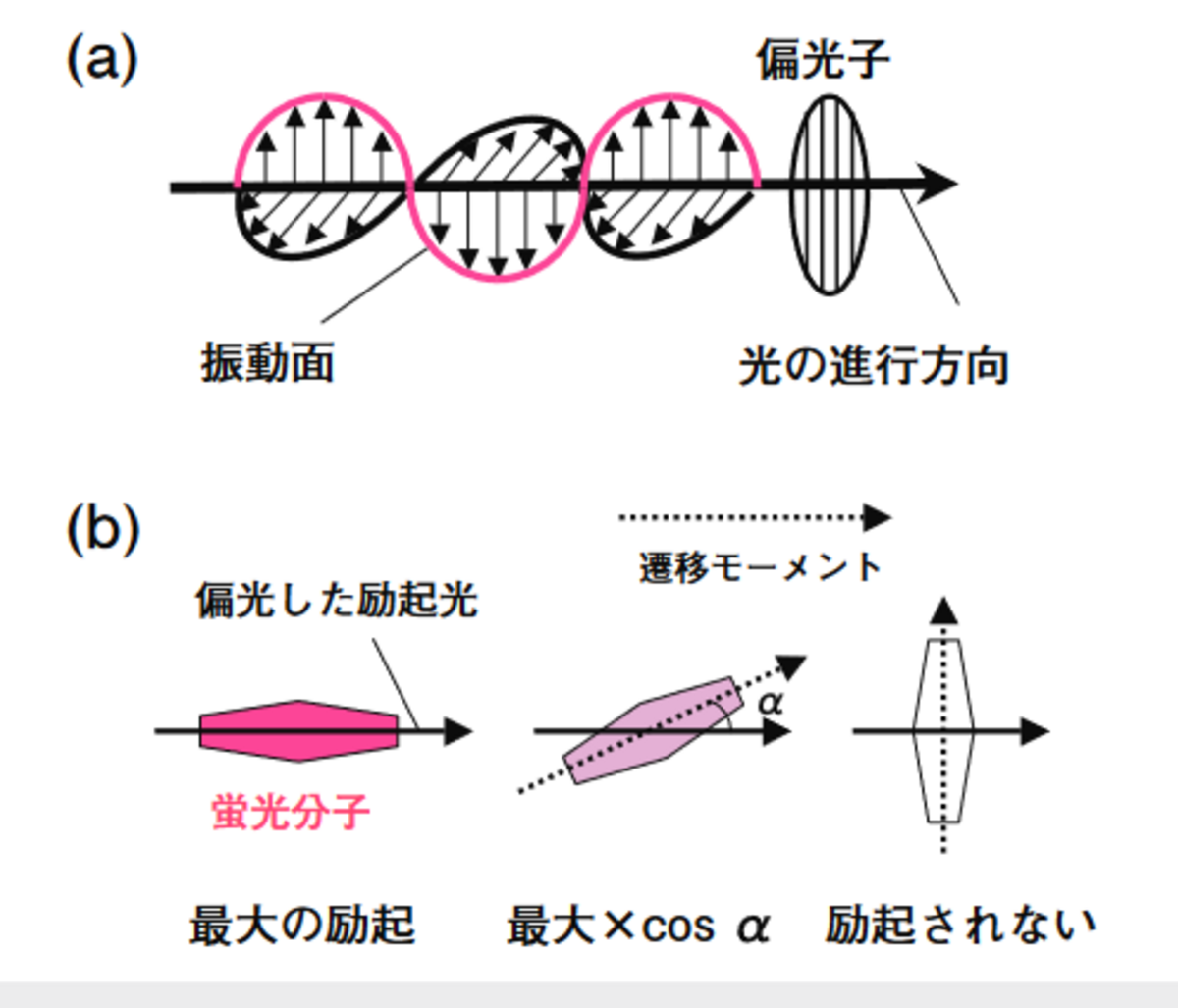
光は横波の性質を持っており,進行方向に対して垂直な平面内で振動している(図2(a))。一方,蛍光分子が励起される場合,S0→S1遷移には分子構造によって決まる向きがあり(これを遷移モーメントと呼ぶ),直線偏光による入射光の振動方向が遷移モーメントと平行であれば励起効率は最大で,直交していれば0になる(これを光選択という)。従って,例えば水溶液中でランダムに分散している蛍光分子であっても,直線偏光で励起されるのはその時点で遷移モーメントが平行な分子集団に限られる(図2(b))。一方,発する蛍光にも固有の向きがあり,強く偏光していれば入射光と平行な螢光の偏光成分が強くなる。この偏光の度合いを表すのに便利なパラメーターが蛍光異方性r(fluorescence anisotropy)で,式(2)で表される。
![]() (2)
(2)
ここで,I//とI⊥はそれぞれ入射光の偏光方向と平行及び垂直な蛍光の偏光成分である。もし蛍光分子が激しく回転ブラウン運動していれば,I//が小さくなりI⊥が大きくなるので,rは小さくなる。これが偏光解消である。従って,rを調べることで蛍光分子の回転運動の度合いが理解される。さて,蛍光寿命の計測と同様にrの時間変化r(t)を調べると何がわかるのだろう。r(t)は式(3)で表される。
![]() (3)
(3)
ここで,r0は分子運動がない時に期待される異方性,θは回転相関時間(fluorescence rotational correlation time)であり,ナノ秒領域の分子の振る舞いを考える上で最も重要なパラメータである。ここで,θの意味について考えてみたい。分子を剛体球とみなした場合,θは式(4)で表される。
![]() (4)
(4)
ここで,kBはボルツマン定数,Tは絶対温度,vは分子の体積,ηは溶媒の粘度である。従って,θさえ求めることができれば,vとηの積がわかる。vがわかっていれば溶媒の粘度ηが求まるし,ηがわかっていれば分子体積vが求まる。こうした原理を生体膜の研究に応用した例を次に紹介する。
図2 偏光と光選択 (a)光は横波で,進行方向と垂直な面で振動 偏光子を通せば特定の振動面にある光だけを得ることができる(ここでは赤で示す)。 (b)光選択の原理 入射光の振動方向が,分子の遷移モーメントと平行であれば励起効率は最大,直交していれば0である。
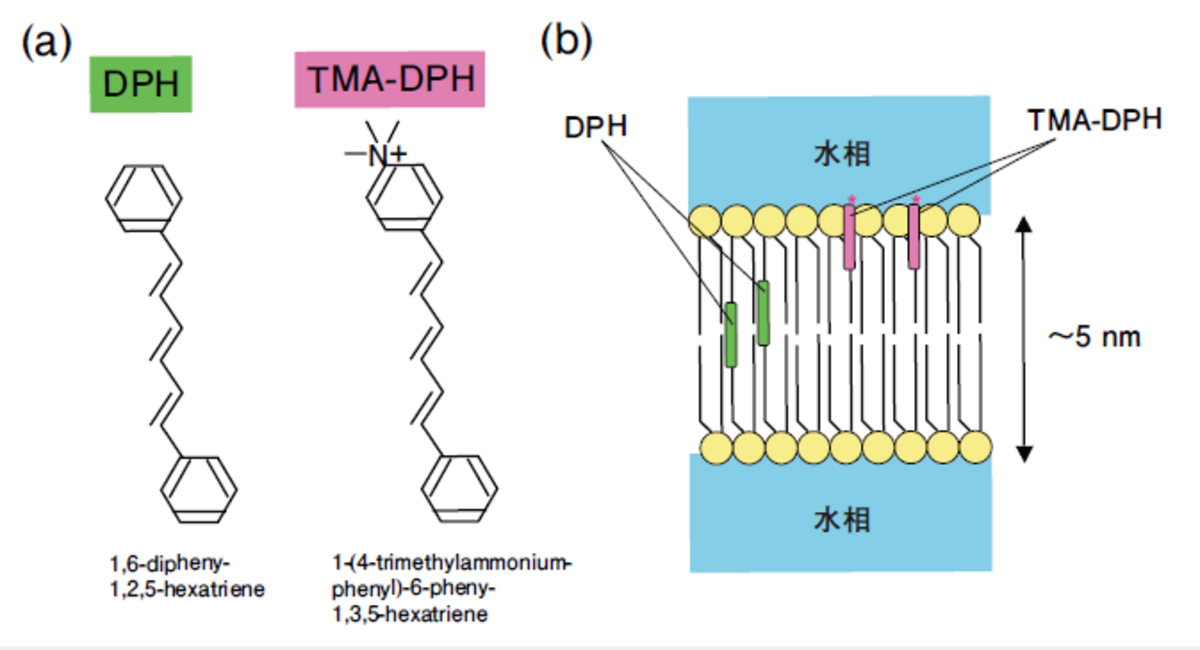
細胞やオルガネラ(細胞内小器官)はリン脂質二重層の膜におおわれ,特定のタンパク質や代謝中間体がコンパートメント化されている。細胞を包み込んでいるのが細胞膜(plasma membrane)で,そこには外界シグナルを伝達する受容体やチャネル,イオン環境を整えるポンプ,アミノ酸などを運び込むトランスポータがぎっしりと敷きつめられている。膜タンパク質の様々な機能を働かせるために肝心なのが,適切な生体膜の状態である。厚さわずか5nmのリン脂質二重層のダイナミクスを知るのによく使われるのが,DPH(1,6-diphenyl-1,3,5-hexatriene)とそのアナログTMA-DPH1-(4-trimethylammonium-phenyl-1,3,-hexatriene)である(図3(a))。
これらの分子は水溶液中では励起しても蛍光を発しないが,脂質中では強い蛍光を発する。また,棒状で強い偏光特性をもつため,図3(b)のように膜に埋め込まれた時,偏光解消から脂質二重層の動的構造がわかる。なお,TMA-DPHは極性基をもつためリン脂質の頭部にアンカーされる。
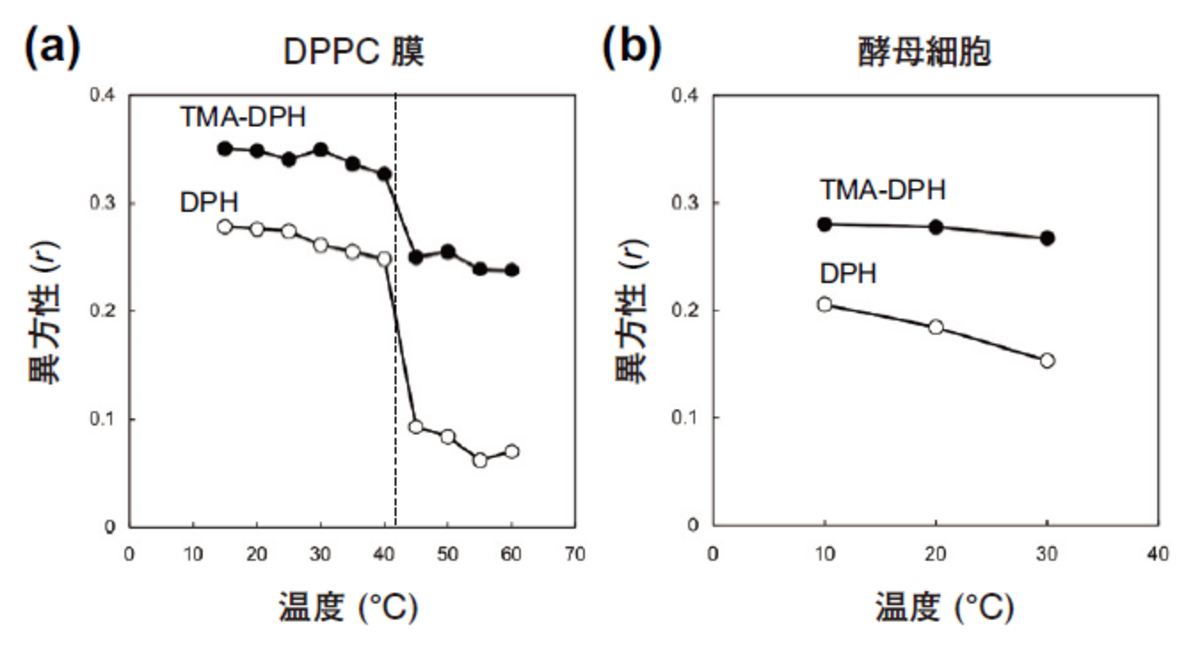
人工膜DPPC(dipalmitoylphospha tidylcholine)をDPHとTMA-DPHでそれぞれラベルし,定常光励起によって異方性r値の温度依存性を調べたのが図4(a)である。DPHのr値は42 ℃付近を境に急激に低下している。42 ℃はDPPCの相転移温度である。この温度以下でDPPC膜はゲルなので,DPHの分子運動が強く束縛されている。ところが,42 ℃を越えると液晶相となり,急速に分子の運動性が増し偏光解消が起こる。その結果としてr値は低下する。一方,TMA-DPHの異方性r値も42 ℃で急激に低下しているが,その度合いはDPHほど顕著ではない。これは,TMA-DPH分子の末端がリン脂質の頭部にアンカーされているせいで,液晶相にあっても分子の運動性がある程度制限されているためである。
これを出芽酵母Saccharomyces cerevisiaeの生きた細胞で実施した例を次に示す。まず,酵母細胞をリン酸緩衝液で洗浄後,5 μMのDPHあるいはTMA-DPHで10分間室温でラベルする。次に余分な試薬を洗い流し,異方性の温度依存性を調べたのが図4(b)である。人工膜と違って相転移に伴う急激なr値の低下は観察されないが,昇温と共に連続的に偏光解消が激しくなっていくのがわかる。一方,TMA-DPHでは温度依存性が小さいことが興味深い。
図3 DPHとTMA-DPAの分子構造と脂質二重層への局在 (a)DPHとTMA-DPAの分子構造 (b) DPHは脂質二重層の中央付近に,TMA-DPAは表面近くに局在する
図4 DPPCと酵母細胞膜におけるDPHとTMA-DPHの異方性測定 (a) DPPC膜にDPHとTMA-DPHを取り込ませ,定常光励起により異方性r値の温度依存性を調べた。点線(42℃)より低温側がゲル相,高温側が液晶相である (b) 酵母細胞膜にDPHとTMA-DPHを取り込ませ,定常光励起により異方性r値の温度依存性を調べた。生体膜では相転移が見られないのがわかる
最近,筆者らは蛍光寿命測定装置FluoroCubeを用いて,酵母細胞膜中のDPHやTMA-DPHの偏光解消を調べ始めた。詳しくは原著論文(投稿準備中)にゆずるとして,以下におおまかな結果を記したい。
定常光励起による前述の測定と同様,細胞をラベルし,異方性の寿命を計測した。レーザ光源にはNanoLED-375Lを用い,460 nmにおける蛍光を測定した。その結果,DPHでは蛍光寿命τそのものは25 ℃と10 ℃とで大差なく約8 nsだった。ところが,回転相関時間θは25 ℃では3 ns(回転が速い),それに対して10 ℃では5 ns(回転が遅い)となり,約1.7倍の差が見られた。このことは,低温によって脂質アルキル鎖の運動性が低下し,DPHの偏光解消が低減することを示している。一方,TMP-DPHでは25 ℃と10 ℃共にτは約14 nsだった。θについては,まだ測定結果にふれが大きいのだが,25 ℃では5~10 ns,10 ℃では15~20 nsという値が得られている。やはり,低温では膜の分子運動の低下が見られる。
FluoroCubeでは非常に広い範囲で多成分解析が可能である。このことは生体試料解析する際に重要である。例えば,細胞膜は人工脂質と違って組成が複雑であり,真核生物では大まかに分けてもグリセロリン脂質,スフィンゴ脂質,ステロール及び膜タンパク質からなり,各々の成分が数種から数万種の物質で構成された混合物である。こうした膜が均一である証拠はなく,むしろ不均一でドメイン構造を作っているという傍証が多い。こうした場合,定常光による解析で得られる異方性r値は各ドメインの平均値だが,FluoroCubeでは多成分解析により個々の成分を定量的に求めることができる。ただし,これはあくまで数学的なフィッティングなので,ドメインの存在については他の生化学的解析の裏付けが必要となろう。
生体膜に関して少し視野を広げてみよう。
医療用薬剤の約50%は膜タンパク質をターゲットとしていて,その研究成果は私たちの健康な暮らしに直結する可能性が高い。
また,コレステロールやスフィンゴ脂質の代謝異常で病気になる例がいくつも知られている。一方,ミトコンドリアや小胞体,あるいはゴルジ体といった膜系は,それぞれエネルギー生産,タンパク質合成並びに膜輸送など,生命維持にとって必須な機能を担っている。もし偏光性が高く,オルガネラ膜に特異的に取り込まれる蛍光試薬が入手できれば,膜の物性を深く理解した上で研究を展開できる。
一方,水溶性タンパク質の回転運動に着目した解析もなされるであろう。タンパク質同士の会合・解離は,細胞骨格形成時や細胞内シグナル伝達系の分子間相互作用において重要である。タンパク質を剛体球と仮定すると,前述の通りθは分子体積vに比例する。2量体を形成すればθは2倍に,4量体なら4倍に近い値を示す。蛍光標識したタンパク質は溶液中でどのようなダイナミックな挙動を示すのか?θを求めることでその真の姿が明らかになるに違いない。ライフサイエンスの広範な領域で,蛍光寿命測定や偏光解消法が真価を発揮する日はすぐそこまで来ている。
[1]Joseph R. Lakowicz, Principles of Fluorescence Spectroscopy, 2nd. Ed., Kluwer Academic/Plenum Publishers (1999).
[2]Bernard Valeur, Molecular Fluorescence, Wiley-VCH (2001).
[3]木下一彦・御橋廣眞編,螢光測定―生物科学への応用,学会出版センター (1983)
HORIBAでは、技術情報誌としてReadoutを発行しています。誌名“Readout(リード・アウト)”には、HORIBAが創造・育成した製品や技術に関する情報を広く世にお知らせし、読み取って頂きたいという願いが込められています。